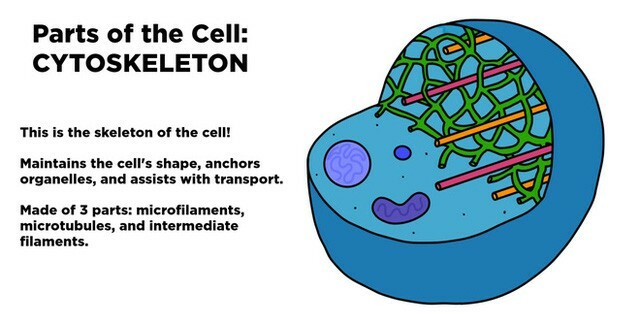
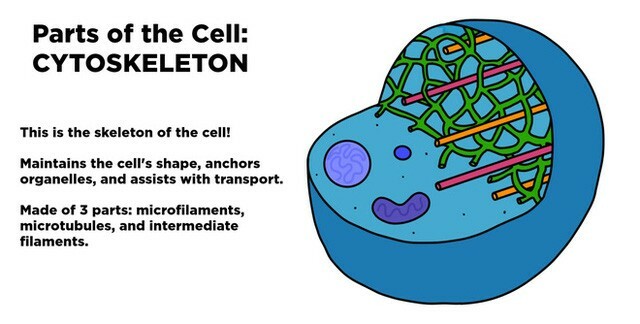
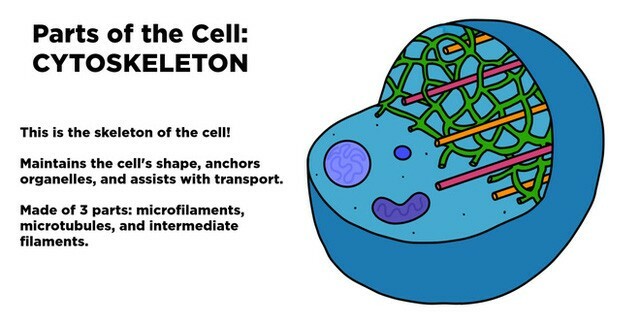
Khung xương tế bào – Vi ống
Không giống như bộ xương trong hệ cơ xương của chúng ta, bộ khung xương tế bào là một cấu trúc động ở mức độ cao, liên tục được tổ chức lại như khi tế bào thay đổi hình thù, khi phân chia, và phản ứng lại môi trường.
Bạn đang xem: Cytoskeleton là gì
Bộ khung xương tế bào không chỉ là “các xương” (bones) của tế bào mà còn là các “cơ” (muscles), và hệ thống khung xương này chịu trách nhiệm trực tiếp đối với những sự vận động trong phạm vi lớn, bao gồm sự bò trườn của tế bào dọc theo một bề mặt, sự co các cơ của tế bào, và những thay đổi trong định hình tế bào diễn ra như khi một phôi phát triển.
Nếu không có bộ khung xương tế bào, các vết thương sẽ không bao giờ lành, các cơ không thể co rút, và tinh trùng cũng không thể đến được với trứng.
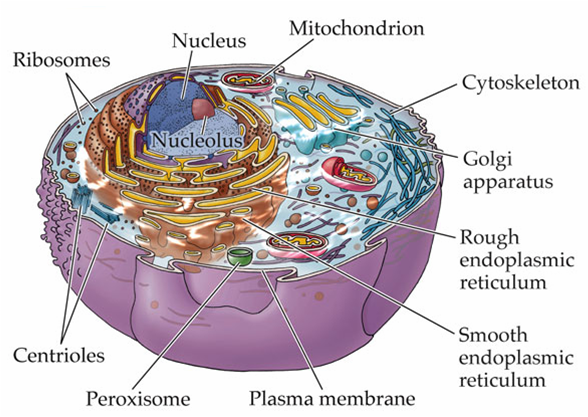
Giống như bất kỳ nhà máy sản xuất một phức hợp sản phẩm, tế bào nhân thực có một hệ thống tổ chức cao bên trong, tại đây các bào quan thực hiện các chức năng chuyên biệt được tập trung tại các khu vực khác nhau và liên kết nhau bởi hệ thống vận chuyển.
Bộ khung xương tế bào kiểm soát vị trí của các bào quan và cung cấp máy móc để vận chuyển giữa chúng. Nó cũng chịu trách nhiệm đối với sự phân ly của các NST thành hai tế bào con trong quá trình phân bào và đối với việc thắt lớp màng tế bào để phân chia tế bào chất của hai tế bào này.
Tế bào động vật với vi ống (xanh lá), các sợi actin (màu đỏ). Những vùng mà hai loại sợi này chồng lên nhau (màu vàng). DNA trong nhân (được thể hiện bằng màu xanh dương).
Bộ khung xương được cấu trúc trên nền tảng là ba loại protein sợi:
sợi trung gian (intermediate filaments)vi ống (microtubules)vi sợi (actin filaments)
Mỗi loại sợi có những tính chất cấu trúc riêng biệt và được hình thành từ các tiểu đơn vị protein khác nhau.
Họ protein sợi cấu tạo nên các sợi trung gian, tiểu đơn vị dạng hình cầu tubulin cấu tạo nên vi ống và các tiểu đơn vị actin hình cầu cấu tạo nên các sợi actin. Trong mỗi trường hợp, hàng ngàn tiểu phần tập hợp lại thành các sợi mảnh chạy dài xuyên suốt tế bào.
1. VI ỐNG
Vi ống có vai trò tổ chức rất quan trọng trong tế bào.
Những ống protein rỗng dài và tương đối cứng có thể nhanh chóng tháo rời ở một vị trí này hay cũng có thể tập hợp lại ở một nơi khác.
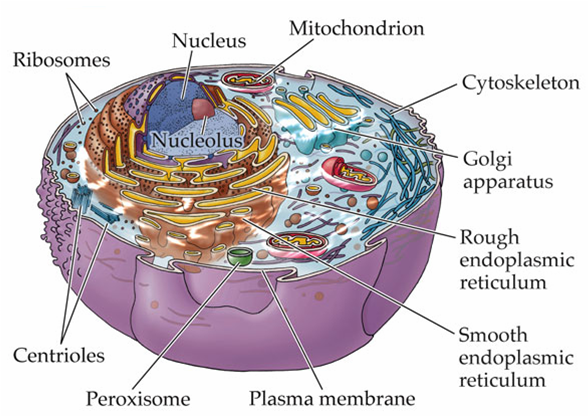
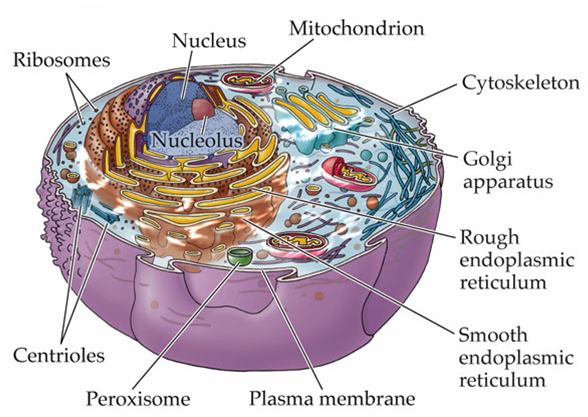
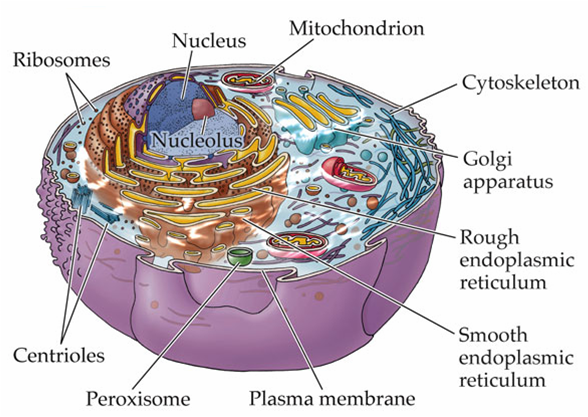
Trong một tế bào động vật điển hình, vi ống phát triển từ một cấu trúc nhỏ gần trung tâm của tế bào được gọi là trung thể (centrosome, hình A, B).
Sự kéo dài ra hướng tới vùng ngoại vi của tế bào nên một hệ thống đường đi trong tế bào, dọc theo các bóng tiết, các bào quan và các ngăn khác của tế bào.
Các vi ống là những phần của hệ thống khung xương chủ yếu chịu trách nhiệm trong vận chuyển và định vị các bào quan có màng trong tế bào và chỉ dẫn sự vận chuyển nhiều đại phân tử thuộc tế bào chất.
Khi một tế bào bước vào nguyên phân, các vi ống tháo rời ra và sau đó lại tập hợp lại thành một cấu trúc rắc rối được gọi là Thoi phân bào (mitotic spindle).
Thoi phân bào cung cấp một bộ máy để phân chia đồng đều các NST cho hai tế bào con trước sự phân chia của tế (hình C).
Vi ống cũng có thể hình thành những cấu trúc ổn định, ví như của lông rung (cilia) và roi (flagella) (hình D); những cấu trúc dạng lông như thế này chạy từ bề mặt từ nhiều tế bào nhân thực, vừa sử dụng chúng để “bơi” vừa để “xua” dòng chất lỏng trên bề mặt của nó. (Roi của vi khuẩn có một cấu trúc khác hoàn toàn và cho phép các tế bào bơi cũng với cơ chế khác).
Vi ống là những ống rống với cấu trúc đặc biệt
Các vi ống được xây dựng từ các đơn vị – các phân tử tubulin – mỗi phân tử là một nhị hợp gồm có 2 protein cầu rất giống nhau được gọi là α-tubulin và β-tubulin và bám chặt lấy nhau bởi những tương tác không cộng hóa trị.
α & β tubulins, trọng lượng phân tử 55 kDa, tương đồng về cấu trúc.
Nhị hợp tubulin xếp lại với để tạo nên thành của một cấu trúc vi ống rỗng hình trụ.
Dạng ống này được tạo nên bởi 13 sợi nguyên (protofilaments) song song nhau, mỗi một chuỗi sợi của tubulin nhị trùng dựa vào α- và β – tubulin suốt dọc chiều dài của nó.
Mỗi sợi nguyên có cấu trúc phân cực, với một đầu tận cùng là α – tubulin và đầu còn lại là β – tubulin, và sự phân cực này – cho thấy tính định hướng trong cấu trúc – giống nhau ở tất cả các sợi nguyên, giúp tạo nên tính phân cực của cả một vi ống.
Đầu β – tubulin là đầu dương, đầu α – tubulin là đầu âm.
Cấu tạo vi ống và ảnh hiển vi chụp vi ống theo các hướng
Trong các dung dịch tinh khiết của tubulin trong ống nghiệm, các nhị hợp tubulin sẽ thêm vào cả hai đầu đang phát triển của vi ống.
Tuy nhiên chúng thêm vào đầu (+) nhanh hơn là đầu (–), đây cũng là lý do vì sao các đầu được đặt tên như thế chứ không phải vì sự tích điện.
Sự phân cực của vi ống – thực tế rằng cấu trúc của nó có định hướng rõ ràng, với hai đầu khác nhau về phương diện hóa học và chức năng – rất quan trọng, cả cho việc hình thành các vi ống và vai trò của nó ngay khi nó được hình thành.
Nếu vi ống không phân cực, nó có thể đã không hướng dẫn được quá trình vận chuyển nội bào chẳng hạn.
Trung thể là trung tâm hình thành vi ống của tế bào động vật
Trong các tế bào, các vi ống tỏa ra từ một trung tâm cấu trúc chuyên biệt, nó kiểm soát vị trí, số lượng, và sự đinh hướng của các vi ống.
Trong tế bào động vật, lấy ví dụ trung thể (centrosome hay MTOC – trung tâm tổ chức vi ống) – thường gần sát nhân khi tế bào không phân chia – tổ chức các vi ống tỏa ra xung quanh khắp tế bào chất.
Trung thể bao gồm một đôi trung tử (centrioles), bao quanh bởi một mạng lưới các protein (chất quanh trung tử).
Trung thể bao gồm hàng trăm cấu trúc dạng vòng tròn được hình thành từ một dạng đặc biệt của tubulin – được gọi là γ – tubulin, và mỗi phức hệ vòng tròn γ – tubulin có vai trò như một điểm bắt đầu, hay vị trí phóng xạ (nucleation site) đối với sự phát triển của một vi ống.
Trung tâm phát triển vi ống (MOTC), ví dụ trung thể ở tế bào động vật
Nhị hợp αβ – tubulin thêm vào mỗi vòng γ – tubulin trong một sự định hướng đặc trưng, với hệ quả là đầu (–) của mỗi vi ống được gắn vào trung thể còn sự phát triển diễn ra chỉ ở đầu (+) rồi kéo dài trong tế bào chất.
Cặp trung tử ở trung tâm của một trung thể (tế bào động vật) có cấu trúc kỳ lạ; mỗi trung tử đặt vuông góc với cái còn lại, tạo nên từ một khối hình trụ của các vi ống ngắn.
Các trung tử không có vai trò gì trong việc tỏa ra của vi ống tại trung thể (chỉ một mình phức hệ vòng γ-tubulin là đủ rồi), và chức năng của chúng vẫn còn là một vài bí ẩn, đặc biệt là ở các tế bào thực vật không có chúng.
Xem thêm: Corporation Là Gì – Joint Stock Company, Corporation Hay Group
Trung tử hoạt động như là trung tâm tổ chức các vi ống trong lông mao và roi (tiên mao và tiêm mao), ở đây chúng được gọi là “các thể gốc” (basal bodies).
Sự phát triển vi ống biểu thị tính năng động bất thường (dynamic instability)
Ngay khi một vi ống bắt đầu được cấu tạo từ vòng γ, nó phát triển ra xa từ trung tâm tổ chức trong nhiều phút nhờ vào quá trình thêm nhị hợp tubulin vào đầu dương (+).
Sau đó, không hề có một sự báo hiệu nào, vi ống có thể bất thình lình chịu một sự biến đổi làm cho chúng co lại rất nhanh về phía trung tâm do sự mất các nhị hợp tubulin từ đầu (+) của nó. Nó có thể co rút một phần và sau đó cũng không kém bất ngờ, tiếp tục phát triển dài ra, hoặc nó có khi biến mất hoàn toàn, bị thay thế bởi một vi ống mới phát triển từ chính vòng γ-tubulin đó.
Đặc tính đáng chú ý này – sự chuyển đổi qua lại giữa quá trình trùng hợp và giải trùng hợp – được xem là “tính năng động thất thường” (dynamic instability). Nó cho phép các vi ống chịu được quá trình tái cấu trúc nhanh, và quyết định chức năng của nó.
Trong một tế bào bình thường, trung thể (hay các trung tâm tổ chức khác) liên tục “phóng” ra các vi ống mới từ các hướng khác nhau trong một dạng như thăm dò, nhiều trong số chúng sau đó lại rụt vào.
Một vi ống phát triển từ trung thể dù thế nào vẫn có thể được tránh khỏi quá trình tháo rỡ nếu đầu (+) của nó được làm cho bền vững bởi được đính kèm với các phân tử khác hoặc được tế bào cấu trúc để ngăn cản sự giải trùng hợp của nó.
Nếu được ổn định bằng cách gắn kèm với một cấu trúc ở 1 vùng cách xa của các tế bào, các vi ống sẽ thiết lập một liên kết tương đối ổn định giữa cấu trúc này và trung thể.
Đầu (+) ban đầu có thể tự do nhưng có thể được ổn định hóa nhờ vào việc gắn với các protein đặc hiệu (A),các vi ống mới được hình thành từ trung tử từ nhiều hướng khác nhau trước khi co lại một cách ngẫu nhiên. Nếu đầu (+) bắt gặp một protein ( mũ protein hay capping protein màu hồng) tại một vùng đặc trưng trên vỏ (màng) tế bào, nó sẽ được ổn định hóa (B). Cuối cùng sẽ chuyển tế bào thành một dạng phân cực mạnh (D).
Điều hòa hoạt động của các vi ống bằng các thuốc
Thuốc có thể ngăn cản quá trình trùng hợp hay giải trùng hợp của các nhị hợp tubulin có tác dụng nhanh và sâu sắc trong sự tổ chức của vi ống. Được xem là thoi tơ vô sắc, vi ống hướng dẫn các NST trong suốt quá trình phân bào.
Nếu một tế bào đang trong kỳ phân bào mà bị tiếp xúc với colchicine, thứ có thể bám chặt vào các nhị hợp tubulin tự do làm ngăn cản quá trình trùng hợp của chúng thành vi ống. Thoi phân bào nhanh chóng biến mất, và tế bào không thể phân chia các NST về hai phía.
Taxol thì có cơ chế tác động ngược lại. Nó bám chặt vào vi ống và ngăn không cho chúng tháo rỡ các đơn vị tubulin. Vì các đơn vị mới vẫn có thể thêm vào, vi ống cứ phát triển chứ không thể co rút.
Tuy nhiên chúng lại gây ra ảnh hưởng như colchicine – làm ngừng quá trình phân chia tế bào trong kỳ phân bào. Các thí nghiệm chỉ ra rằng để thoi tơ thực hiện chức năng, thì các vi ống phải có khả năng tập hợp hay phân giải.
Các vi ống và các protein vận chuyển định vị các bào quan trong tế bào chất
Các vi ống và các motor protein thể hiện một vai trò quan trọng trong việc định vị các bào quan bên trong một tế bào nhân thực.
Trong hầu hết các tế bào động vật, các vi ống của lưới nội chất vươn tới hầu hết các giới hạn (vùng biên) của tế bào. Khi tế bào lớn lên, protein kinesins đính kèm với mặt ngoài của màng lưới nội chất (thông qua các protein thụ thể) lôi kéo lưới nội chất lan ra theo vi ống, kéo căng giãn như một tấm lưới.
Protein dyneins trong tế bào chất đính vào màng của phức hệ Golgi, kéo phức hệ Golgi theo hướng ngược lại, hướng vào nhân. Bằng cách này, các khu vực khác nhau bên trong màng sinh chất – tính quyết định đối với vai trò riêng của chúng – được hình thành và duy trì.
Khi tế bào được xử lý bằng colcicine – một loại thuốc làm cho vi ống tan ra – cả lưới nội chất và bộ máy Golgi đều thay đổi vị trí của nó một cách rõ rệt.
Lưới nội chất (vốn tiếp xúc với màng nhân) thì tan rã xung quanh nhân, bộ máy Golgi (không được đính với bào quan nào) cũng bị phân mảnh thành các túi nhỏ, những túi mà sau đó được phát tán khắp tế bào chất.
Khi mà colcicine được loại đi, các bào quan lại trở về vị trí ban đầu của nó, được kéo lê bởi các motor proteins di chuyển dọc theo các vi ống mới được tái hình thành.
Lông và roi chứa đựng các vi ống bền vững có thể cử động được là nhờ vào dynein
Chúng ta đã sớm đề cập rằng nhiều vi ống trong tế bào được làm bề vững hóa thông qua việc liên kết với các protein và từ đó không xảy ra hiện tượng tính năng động bất thường (dynamic instability).
Tế bào sử dụng các vi ống này như là một giá đỡ vững trong sự dựng xây các cấu trúc phân cực, bao gồm các lông và roi cử động.
Lông rung (Cilia) có cấu trúc dạng lông đường kính khoảng 0.25 μm, được bao bọc bởi màng sinh chất ( tương tự nhưng không phải là màng của tế bào), phát triển ra từ bề mặt của nhiều tế bào nhân thực; mỗi sợi lông bao gồm một lõi vi ống bề vững, được sắp xếp thành một bó, lớn lên từ basal body của bào tương, nó đáp ứng như là một trung tâm tổ chức.
Lông rung cử động theo kiểu quất (như là quất roi ngựa), vừa để xua đẩy chất lỏng trên bề mặt của một tế bào hoặc là đẩy các tế bào đơn đi qua một chất lỏng. Ví dụ ở một số nguyên sinh vật, sử dụng lông để thu nhặt các hạt thức ăn và còn dùng cho sự di chuyển của chúng nữa.
Tại các tế bào biểu mô lớp niêm mạc đường hô hấp của người, một lượng lớn các lông nhu động (hơn một tỉ lông mỗi centimet vuông), quét đi những đám dịch nhày chứa các hạt bụi và các tế bào chết từ họng.
Một cách tương tự, sự nhu động của các lông trên tế bào ở thành ống dẫn trứng tạo ra một dòng lưu chuyển giúp đưa trứng đi vào bên trong ống dẫn trứng tới tử cung. Mỗi “sợi” lông cử động như một mái chèo nhỏ, di chuyển thành một trình tự lặp lại sinh ra sự lưu chuyển của dòng chất lỏng trên bề mặt tế bào.
Roi (flagellum) ở tinh trùng hay nhiều động vật nguyên sinh rất giống với lông rung về cấu trúc bên trong nhưng roi thường dài hơn nhiều. Nó được cấu tạo để vận chuyển cả một tế bào, còn hơn cả việc di chuyển dòng chất lỏng qua bề mặt tế bào. Các roi thường liên tục sinh ra các sóng dọc theo chiều dài của nó, đẩy tế bào đi.
Ảnh chụp liên tiếp sự chuyển động của một tinh trùng
Các vi ống trong lông và roi có khác một chút so với các vi ống trong tế bào chất, chúng được sắp xếp trong một kiểu mẫu tỉ mỉ và đặc biệt, một trong những thứ dễ “đập vào mắt” nhất khi quan sát bằng kính hiển vi điện tử.
Lát cắt ngang lông rung chỉ ra rằng 9 bộ kép vi ống được xếp thành một vòng tròn xung quanh một cặp vi ống đơn. Kiểu “9+2” này là một đặc tính của hầu hết tất cả các tế bào nhân thực có lông và roi – từ những tế bào của các động vật nguyên sinh cho tới con người.
Sự chuyển động các lông hay roi được tạo ra bởi sự uốn cong lõi của nó mỗi khi các vi ống trượt lên nhau.
Các vi ống liên kết với một lượng lớn các protein phụ. Một vài trong số các protein này có vai trò như các liên kết ngang để giữ các bó vi ống; các protein khác cung cấp lực làm cho lông uốn cong.
Protein phụ quan trọng nhất là một motor protein ciliary – dynein, sinh ra lực uốn cong lõi. Nó liên hệ mật thiết tới protien dynein trong tế bào chất và có chức năng rất tương đồng.
Ở người, các khuyết tật di truyền trong ciliary – dynein gây ra hội chứng Kartagener. Người đàn ông mang rối loạn này thì vô sinh bởi vì tinh trùng của họ không cử động và ai bị hội chứng sẽ tăng tính nhạy cảm đối với tình trạng nhiễm trùng đường phế quản bởi vì lớp nhu mao trong niêm mạc hô hấp bị tê liệt và vì thế không có khả năng loại bỏ vi khuẩn hay những mảnh vụn khỏi phổi.
Xem thêm: Cmm Là Gì – Mấy Bạn Cho Mình Hỏi Vậy
Bài viết tham khảo từ sách Essential cell biology Fourth Edition By Bruce Alberts, Dennis Bray, Karen Hopkin, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter.
Chuyên mục: Hỏi Đáp