Hoạt động của nhiều con đường trao đổi chất có thể được điều hoà nhờ việc điều chỉnh hoạt tí;nh của các enzyme điều chỉnh. Mục này mô tả các enzyme nói trên và đề cập vai trò của chúng trong việc điều chỉnh hoạt tí;nh của con đường.
Bạn đang xem: Allosteric là gì
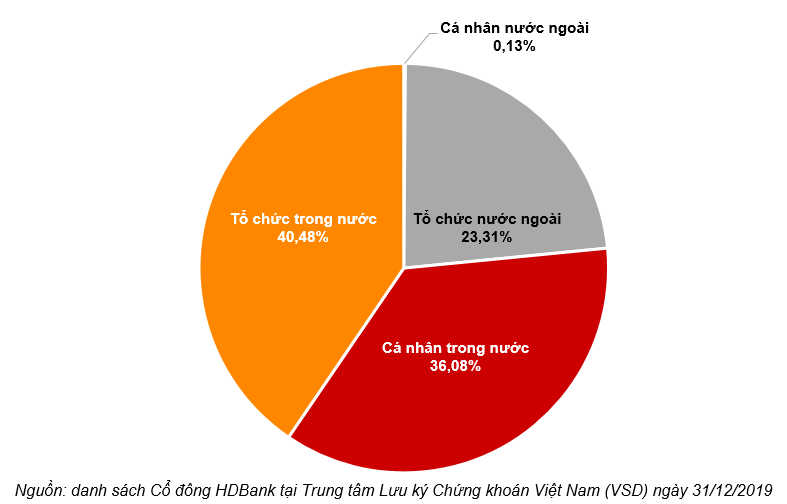
Điều chỉnh dị lập thể
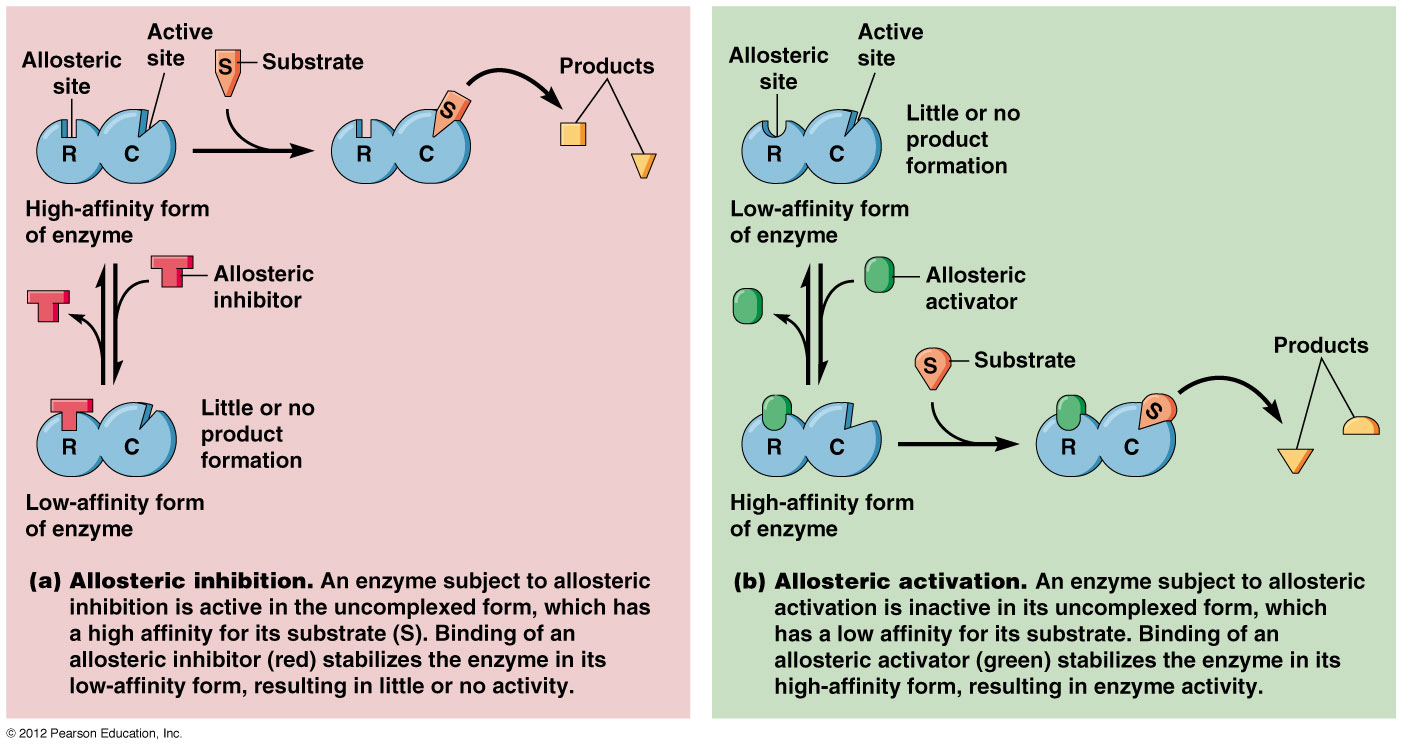
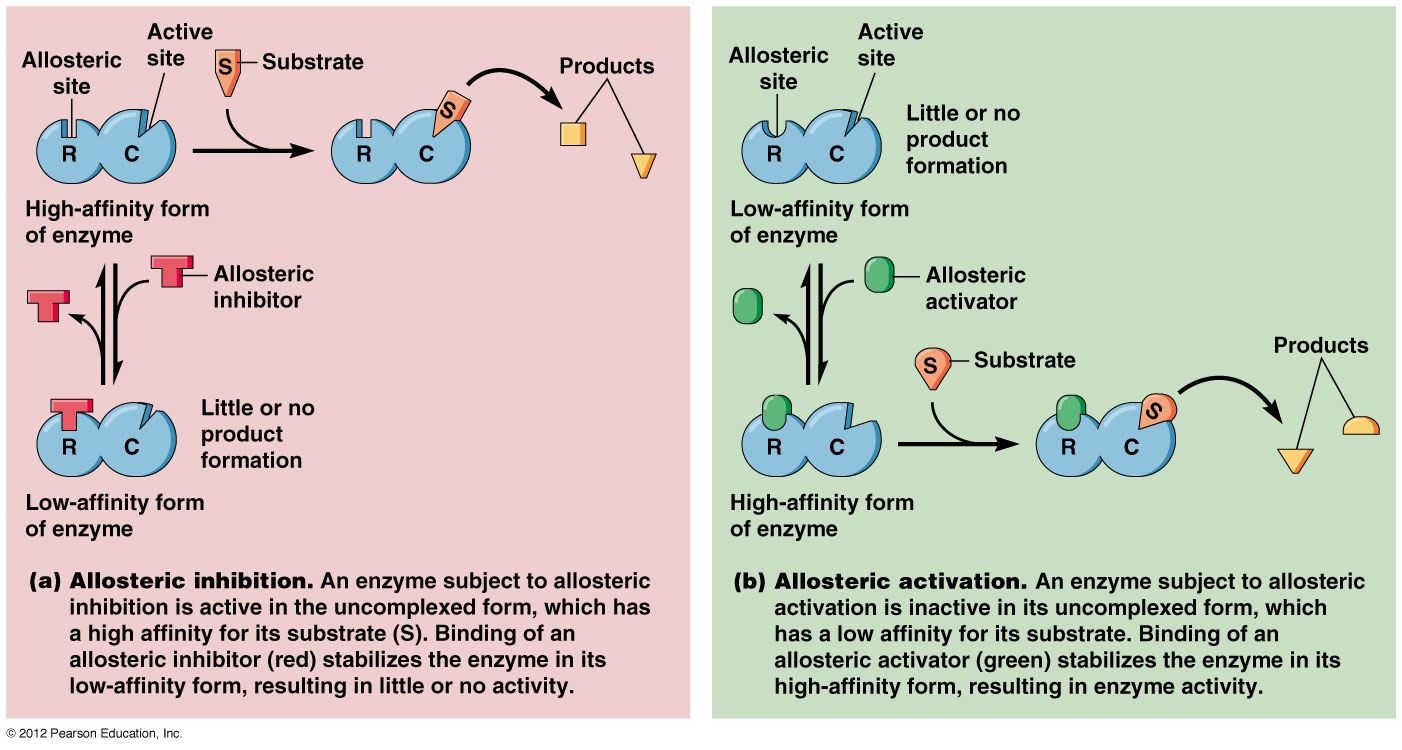
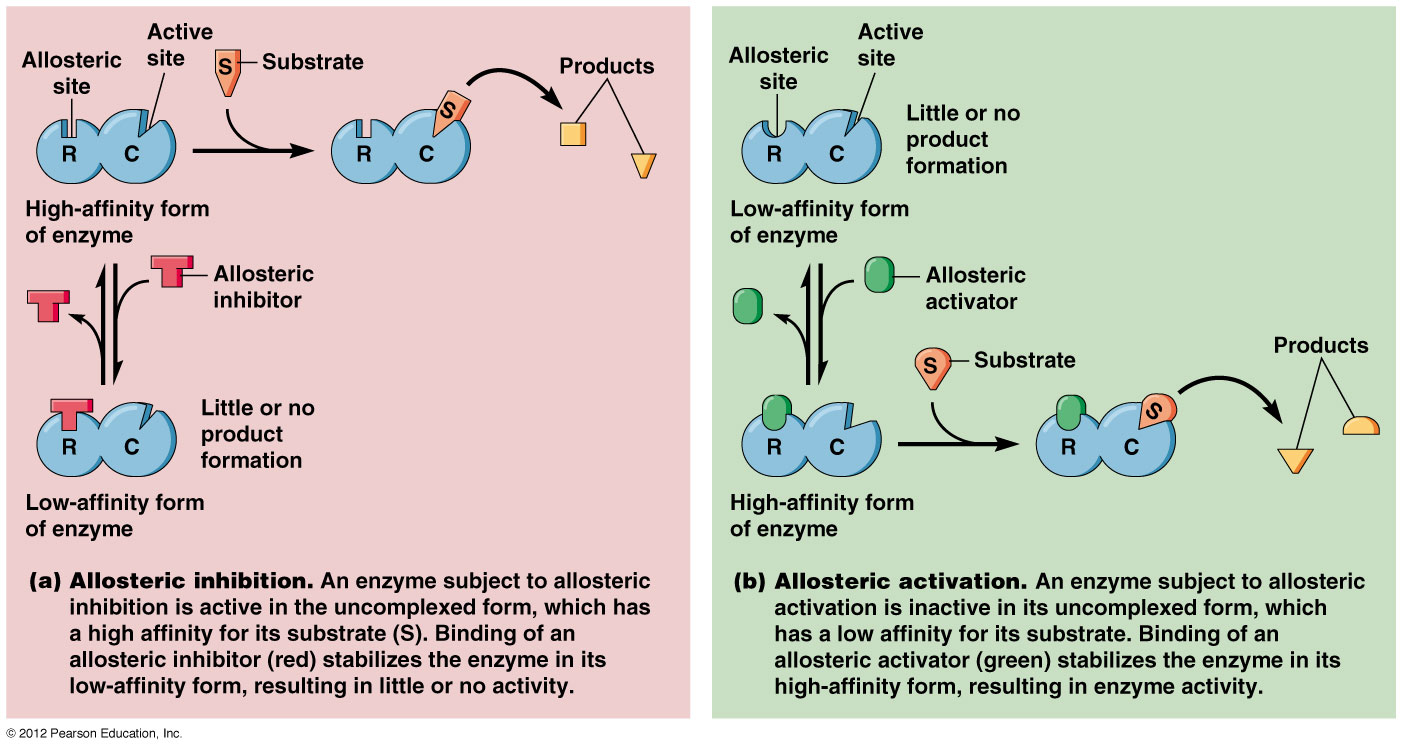
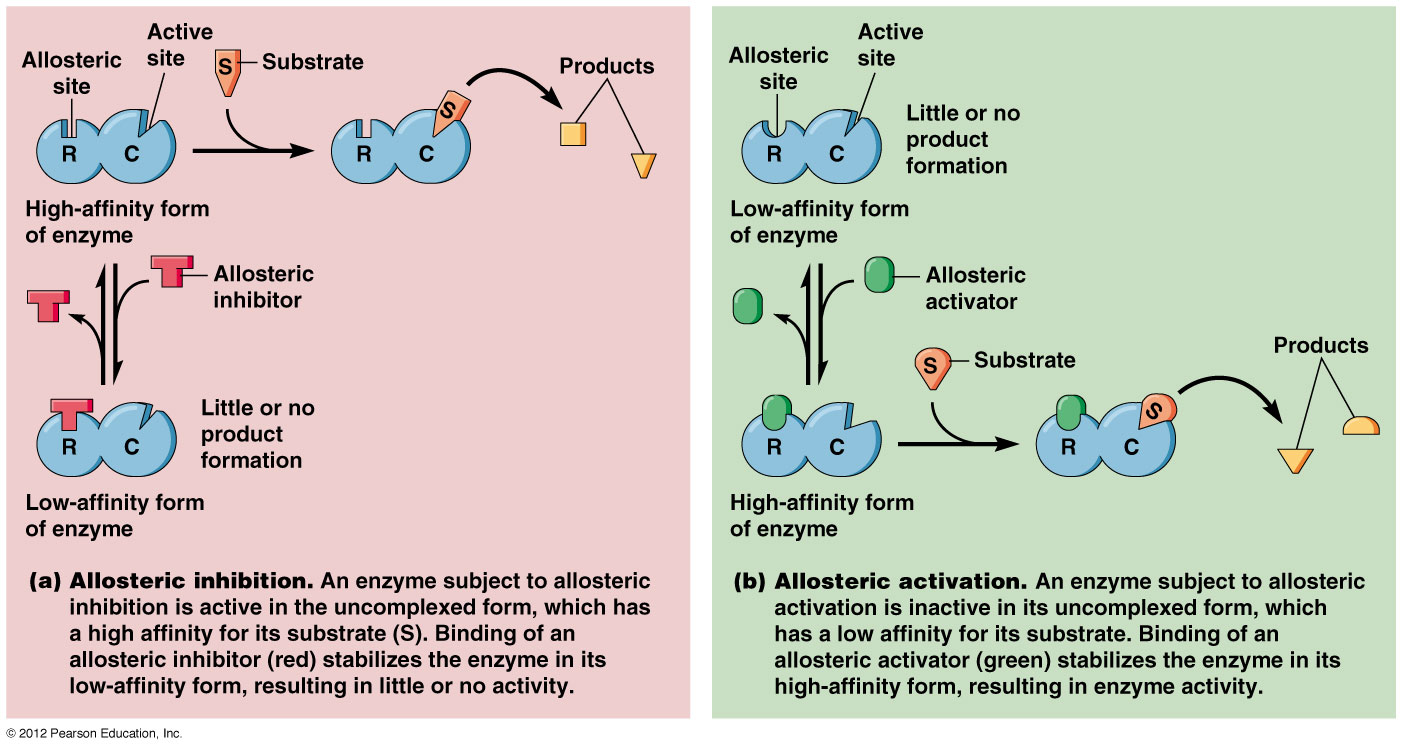
Các enzyme điều chỉnh thường là các enzyme dị lập thể (allosteric enzymes). Hoạt tí;nh của một enzyme dị lập thể bị thay đổi bởi một phân tử nhỏ gọi là effector (effector, chất tác động) hoặc modulator (modulator, chất điều biến). Effector liên kết thuận nghịch nhờ lực không – cộng hoá trị vào một vị trí; điều chỉnh (regulatory site) tách biệt khỏi vị trí; xúc tác (catalytic site) và gây ra sự thay đổi trong hình dạng hoặc hình thể của enzyme (Hình 16.21). Hoạt tí;nh của vị trí; xúc tác do đó bị thay đổi. Một effector dương làm tăng hoạt tí;nh enzyme, một effector âm, trái lại, làm giảm hoạt tí;nh hoặc kìm hãm enzyme. Những thay đổi như vậy trong hoạt tí;nh thường bắt nguồn từ những biến đổi trong ái lực biểu kiến của enzyme đối với cơ chất, tuy nhiên những thay đổi trong tốc độ cực đại cũng có thể diễn ra.
Điều chỉnh dị lập thể
Cấu trúc và chức năng của 1 enzyme dị lập thể. Trong hình bên effector hoặc modulator (chất điều biến) trước hết gắn vào 1 vị trí; điều hòa tách biệt và làm thay đổi hình thể enzyme dẫn đến sự thay đổi hình dạng của vị trí; hoạt động. Vị trí; hoạt động giờ có thể liên kết cơ chất hiệu quả hơn. Ở đây effector là dương tí;nh vì nó kí;ch thí;ch sự liên kết cơ chất và hoạt tí;nh xúc tác. (Theo Prescott, Harley và Klein, 2005)
Các enzyme điều chỉnh thường là các enzyme dị lập thể (allosteric enzymes). Hoạt tí;nh của một enzyme dị lập thể bị thay đổi bởi một phân tử nhỏ gọi là effector (effector, chất tác động) hoặc modulator (modulator, chất điều biến). Effector liên kết thuận nghịch nhờ lực không – cộng hoá trị vào một vị trí; điều chỉnh (regulatory site) tách biệt khỏi vị trí; xúc tác (catalytic site) và gây ra sự thay đổi trong hình dạng hoặc hình thể của enzyme (Hình 16.21). Hoạt tí;nh của vị trí; xúc tác do đó bị thay đổi. Một effector dương làm tăng hoạt tí;nh enzyme, một effector âm, trái lại, làm giảm hoạt tí;nh hoặc kìm hãm enzyme. Những thay đổi như vậy trong hoạt tí;nh thường bắt nguồn từ những biến đổi trong ái lực biểu kiến của enzyme đối với cơ chất, tuy nhiên những thay đổi trong tốc độ cực đại cũng có thể diễn ra.
Sự điều chỉnh ACTase
Phản ứng Aspartate – carbamoyltransferase và vai trò của enzyme này trong việc điều chỉnh sinh tổng hợp pyrimidine. Sản phẩm cuối cùng CTP kìm hãm hoạt tí;nh của ACTase (-) còn ATP lại hoạt hóa enzyme (+). Cacbamoyl Phosphate synthetase cũng bị kìm hãm bởi các sản phẩm cuối cùng của con đường như UMP. (Theo Prescott, Harley và Klein, 2005)
Các đặc tí;nh động học của enzyme không – điều chỉnh chứng minh rằng hằng số Michaelis (Km) là nồng độ cơ chất cần cho một enzyme hoạt động ở tốc độ bằng nửa tốc độ cực đại. Hằng số này chỉ ứng dụng cho các đường cong bão hoà cơ chất hyperbole mà không cho các đường cong xí;ch-ma thường gặp với các enzyme dị lập thể (Hình 16.23). Nồng độ cơ chất cần cho một nửa tốc độ cực đại với các enzyme dị lập thể có đường cong cơ chất xí;ch-ma được gọi là giá trị 0,5 hoặc K0,5.
Xem thêm: Stem Là Gì – Giáo Dục
Một trong các enzyme điều chỉnh dị lập thể được nghiên cứu kỹ nhất đó là Aspartate-carbamoyltransferase (ACTase) ở E. coli. Enzyme xúc tác sự ngưng tụ của cacbamoylphosphate với aspartate tạo thành cacbamoylaspartate (Hình 16.22).
ACTase xúc tác phản ứng quyết định tốc độ của con đường sinh tổng hợp pyrimidine ở E. coli. Đường cong cơ chất bão hoà là xí;ch-ma khi nồng độ của một trong hai cơ chất thay đổi (Hình 16.23)
Động học của Aspartate carbamoyltransferase ở E. coli
CTP là 1 effector âm làm tăng giá trị K 0,5 , còn ATP là 1 effector dương, hạ thấp K 0,5 . V max vẫn là hằng số. (Theo Prescott, Harley và Klein, 2005)
Enzyme có trên một vị trí; hoạt động và sự liên kết của một phân tử cơ chất vào một vị trí; hoạt động sẽ kí;ch thí;ch sự liên kết của cơ chất vào các vị trí; khác. Hơn nữa, cytidine triphosphate (CTP), một sản phẩm cuối cùng của sinh tổng hợp pyrimidine, kìm hãm enzyme, trái lại ATP (purine) lại hoạt hoá enzyme. Cả hai effector thay đổi giá trị K0,5 của enzyme nhưng không thay đổi tốc độ cực đại của enzyme. GTP kìm hãm bằng cách nâng cao K0,5 hoặc chuyển dịch đường cong bão hoà cơ chất lên các giá trị cao hơn. Điều này cho phép enzyme hoạt động chậm hơn ở một nồng độ cơ chất đặc biệt khi CTP có mặt. ATP hoạt hoá bằng cách chuyển đường cong tới các giá trị nồng độ cơ chất thấp hơn khiến cho enzyme hoạt động cực đại qua một phạm vi nồng độ cơ chất rộng lớn. Do đó, khi con đường hoạt động tới mức nồng độ CTP tăng quá cao hoạt tí;nh ACTase sẽ giảm và tốc độ tạo thành sản phẩm cuối cùng bị chậm lại. Trái lại, khi nồng độ sản phẩm cuối cùng ATP tăng lên so với CTP, ATP sẽ kí;ch thí;ch tổng hợp CTP thông qua tác dụng lên ACTase.
Xem thêm: Container-fluid Là Gì – Hướng Dẫn Sử Dụng Bootstrap Container
Aspartate carbamoyltransferase ở E. coli cung cấp một ví; dụ rõ rệt về các vị trí; điều chỉnh và vị trí; xúc tác riêng rẽ trong các enzyme dị lập thể. Enzyme là một protein lớn gồm 2 dưới đơn vị xúc tác và 3 dưới đơn vị điều chỉnh (Hình 16.24a). Các dưới đơn vị xúc tác chỉ chứa các vị trí; xúc tác và không chịu ảnh hưởng bởi CTP và ATP. Các dưới đơn vị điều chỉnh không xúc tác phản ứng nhưng có các vị trí; điều chỉnh liên kết CTP và ATP. Khi liên kết vào enzyme hoàn toàn các effector này gây ra những thay đổi về hình thể trong các dưới-đơn vị điều chỉnh, sau đó trong các dưới đơn vị xúc tác và các vị trí; xúc tác. Enzyme có thể thay đổi thuận nghịch giữa một dạng T í;t hoạt động và một dạng R hoạt động hơn (Hình 16.24 b, c). Do đó vị trí; điều chỉnh ảnh hưởng đến vị trí; xúc tác ở khoảng cách khoảng 6,0 nm.
Cấu trúc và điều chỉnh của Aspartate carbamoyltransferase ở E. coli. (Theo Prescott, Harley và Klein, 2005)
Chuyên mục: Hỏi Đáp